Chapitre 4 Expression des besoins des animaux et des apports de la ration
Les animaux se caractérisent par 5 catégories de besoins différents qui doivent être couverts par l’alimentation :
les besoins en énergie,
les besoins matières azotées,
les besoins en minéraux,
les besoins en vitamines et
le besoin en eau.
Les aliments doivent apporter à l’animal tous les constituants nécessaires au bon fonctionnement de l’organisme et à la réalisation des productions qui peuvent être :
du travail,
de la croissance (croît),
de la viande,
un ou des fœtus,
du lait,
des œufs, etc.
L’ajustement des apports alimentaires peut être quotidien mais certaines rations peuvent aussi couvrir les besoins d’une période bien plus longue comme chez les carnivores au zoo. Cette possibilité dépend à la fois de la nature de la dépense et des caractéristiques des animaux.
La détermination des besoins nutritionnels des animaux tient compte de critères tels que :
l’espèce,
la génétique,
le stade physiologique.
Il faut ensuite adapter ces besoins en fonction des performances obtenues.
4.1 Nature des dépenses
4.1.1 Les dépenses d’entretien
Les activités de l’organisme animal sont à l’origine d’une dépense en éléments nutritifs. Les dépenses d’entretien correspondent à la possibilité pour l’animal de se maintenir en vie, sans variation de poids et sans production d’aucune nature. Ce sont :
d’une part, les dépenses dues au fonctionnement de base de l’organisme, et
d’autre part, les dépenses supplémentaires liées aux conditions de vie (station debout, en déplacement, …).
4.1.2 Les dépenses de production
Les dépenses de production correspondent à la réalisation d’une production donnée par l’animal, qui peut être : une croissance, un engraissement, une gestation, la lactation (production laitière), la ponte, un travail (traction bovine).
4.1.3 Application à la vache laitière productrice de lait
Le lait synthétisé dans la mamelle contient en moyenne par kg :
871 g d’eau,
129 g de matière sèche comprenant 48 g de lactose (glucide), 40 g de matières grasses (lipides),
33,5 g de matières azotées,
7,5 g de matières minérales, dont 1,25 g de calcium,
0,95 g de phosphore,
et des vitamines et substances diverses à l’état de traces.
Pour cela, la vache a effectué des dépenses en matériaux et en énergie (en d’autres termes, elle a utilisé des produits finaux de la digestion), expliqué en ce qui suit :
4.1.3.1 Dépense en matériaux
Les constituants organiques du lait sont, dans leur quasi-totalité, synthétisés par la mamelle à partir d’éléments prélevés dans le sang : glucose, acétate, acides gras longs, acides aminés, etc. Pour être plus précis :
le lactose est synthétisé à partir du glucose prélevé dans le sanguin ;
les protéines du lait (caséines, lactoglobulines, lactalbumines) sont synthétisées à partir des acides aminés ;
les graisses du lait sont élaborées en partie à partir des AG longs véhiculés par le sang et inclus sans modification importante dans la matière grasse du lait, et en partie à partir de l’acétate et du β-hydroxybutyrate provenant directement ou indirectement des fermentations du rumen. Dans ce dernier cas, la mamelle réalise des synthèses qui conduisent à la production d’AG à chaînes plus courtes (8 à 14 atomes de carbone).
Les minéraux et les vitamines sont extraits du sang et inclus dans le lait sans transformation ni synthèse.
La production de lait occasionne ainsi une dépense en matériaux (utilisation de nutriments) tels que du glucose, des acides aminés, des acides gras, des minéraux, et des vitamines.
4.1.3.2 Dépense en énergie
Les différentes synthèses nécessaires pour l’élaboration du lait ont lieu dans les cellules sécrétrices des acini. Ces synthèses sont réalisées par des réactions biochimiques complexes. On peut citer, par exemple, la synthèse du lactose à partir d’une molécule de galactose et d’une molécule de glucose (équation 4.1) :
\[Galactose + Glucose ===> Lactose + Eau\]
Le sang ne contenant pas de galactose, les cellules sécrétrices doivent donc au préalable assurer la synthèse du galactose à partir d’une molécule de glucose.
Ces synthèses sont, sous la dépendance d’hormones catalysées par des enzymes, elles ne peuvent avoir lieu sans énergie. En effet, toute activité cellulaire s’accompagne d’une consommation d’énergie provenant de la combustion d’un élément (ou constituant) organique (constituant de la matière organique). Ainsi, la réaction de synthèse précédente (équation 3.1) peut être réalisée par les cellules sécrétrices grâce à l’énergie libérée lors de la combustion d’une molécule de glucose. Globalement, cette réaction de combustion peut s’écrire (équation 4.2) :
\[C_6H_{12}O_6 + 6 O_2 ====> 6 CO_2 + 6 H_2O + énergie\]
En fait, au niveau cellulaire, la réaction de combustion (réaction d’oxydation) est fragmentée en étapes successives. Cela permet une lente libération de l’énergie. La fourniture de l’oxygène nécessaire à la réaction d’oxydation et l’élimination des produits de déchets (\(CO_2\) et \(H_2O\)) sont assurés par la respiration.
L’énergie nécessaire peut provenir d’un élément (constituant) glucidique, lipidique ou protidique, qui peuvent en effet subir une oxydation au niveau cellulaire. Aux dépenses en matériaux s’ajoute donc une dépense en énergie.
Ainsi, les dépenses d’entretien entraînent :
une dépense en matériaux, liée au perpétuel renouvellement des tissus de l’organisme, dont la vitesse varie avec le tissu considéré et avec l’âge. Par exemple, le renouvellement des protéines musculaires se traduit par une perte d’azote sous forme d’urée chez les mammifères et d’acide urique chez les oiseaux, compensée par une synthèse protéique à partir des acides aminés sanguins,
une dépense énergétique liée au maintien des processus vitaux : respiration, circulation, sécrétions internes, tonus musculaire, pousse des poils, activité physique minimale (station debout, déplacements, …), ingestion et digestion de la ration, énergie nécessaire aux synthèses assurant le renouvellement des tissus de l’organisme, etc.
4.2 Besoins et apports alimentaires
4.2.1 Expression des besoins
Alimenter rationnellement les animaux consiste à compenser les dépenses d’entretien et de production grâce à un apport par les aliments, sans carence ni excès. Il s’agit donc de fournir à l’animal un apport nutritif équilibré et en quantité suffisante.
Les éléments nutritifs contenus dans les aliments ne sont pas entièrement disponibles pour la couverture de ces dépenses. En effet, les aliments sont transformés en éléments utilisables en deux étapes principales (Figure 4.1) :
la digestion dont les produits finaux sont absorbés et déversés dans le sang et la lymphe sous forme de nutriments. Le coefficient d’utilisation digestive (CUD) aussi appelé la digestibilité caractérise l’efficacité de cette étape
le métabolisme qui, à partir des nutriments, permet de satisfaire les besoins avec un rendement appelé coefficient d’utilisation métabolique (CUM).
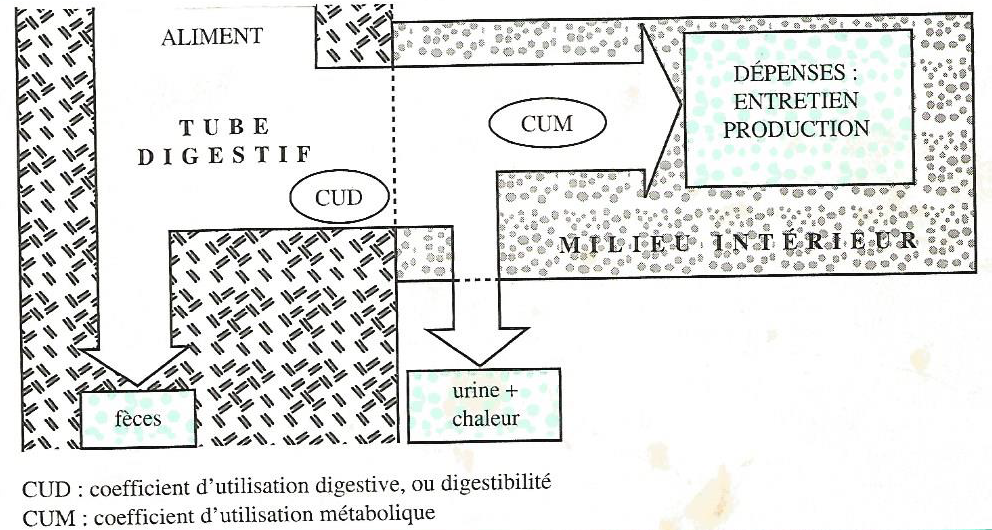
Figure 4.1 Représentation schématique du devenir des apports alimentaires
Il est donc théoriquement possible d’exprimer les besoins à l’un des 3 niveaux suivants :
- celui de l’aliment (composition brute) ;
- celui de l’absorption intestinale (utilization digestive) ;
- celui de la dépense elle-même, c’est-à-dire le niveau cellulaire (utilisation nette).
Le niveau retenu est variable selon la nature de la dépense et l’espèce animale considérées. Dans tous les cas, pour la cohérence du système retenu, la valeur nutritive des aliments doit évidemment être exprimée au même niveau que les besoins des animaux.
4.2.1.1 Chez les ruminants
Le niveau retenu pour l’énergie est celui de la dépense elle-même. Les besoins énergétiques et la valeur énergétique des aliments sont donc exprimés en énergie nette (EN). Pour les matières azotées, le système des PDI (protéines digestibles dans l’intestin) retient le niveau de l’absorption intestinale.
4.2.1.2 Chez le porc et la volaille
C’est le niveau de l’absorption intestinale qui est choisi pour exprimer les valeurs des aliments et des besoins en énergie digestible (ED). Pour les matières azotées, les besoins et les valeurs des aliments sont exprimées en matières azotées totales (MAT) et en acides aminés (AA) au niveau de l’aliment.
Pour les minéraux, on retient dans tous les cas (ruminants et monogastriques), le niveau de l’aliment.
4.2.2 Méthodes d’estimation des dépenses et des apports alimentaires
Deux approches différentes peuvent être utilisées pour estimer les dépenses et les besoins :
la méthode factorielle qui consiste à déterminer le besoin total en sommant les différents besoins partiels correspondant aux différentes fonctions physiologiques. Elle est en principe la plus précise mais sa principale limite réside dans la difficulté de determiner certains besoins partiels, par exemple, le besoin pour la croissance est difficile à définir car on connaît souvent mal la composition du gain de poids.
la méthode globale qui repose sur l’évaluation globale du besoin total à partir d’essais d’alimentation. Elle n’est utilisée que lorsque la méthode factorielle est inutilisable ou trop imprecise.
Les besoins doivent être couverts par les apports alimentaires. Théoriquement les apports alimentaires doivent être strictement égaux aux besoins. En pratique, et en général, on ajuste quotidiennement les apports aux besoins en considérant une marge de sécurité suffisante (très variable selon la nature des besoins), ce qui conduit aux apports alimentaires recommandés. Ainsi donc, les apports alimentaires recommandés (ou recommandations alimentaires) sont supérieurs aux besoins. Les principales raisons justifiant l’adoption d’une marge de sécurité sont :
les incertitudes liées aux caractéristiques des aliments et des matières premières dues à la variabilité de leur composition et/ou à l’imprécision des méthodes d’analyse,
les incertitudes sur la valeur exacte des besoins : l’alimentation se raisonne de plus en plus au niveau d’un troupeau et l’hétérogénéité des individus, en particulier quant à leurs performances, nécessite de prendre des marges de sécurité suffisantes. Toute homogénéisation des performances, quelle qu’en soit la cause, conduit à diminuer les apports recommandés, pour une même performance moyenne,
les incertitudes sur la prévision des quantités consommées dont les causes multiples peuvent être liées à l’animal, à l’aliment, ou au milieu.
Dans certains cas particuliers, les apports alimentaires recommandés peuvent être inférieurs aux besoins car on admet une utilisation des réserves corporelles. Deux situations peuvent être rencontrées :
Animaux à besoins modérés pour lesquels, pour des raisons économiques, on a intérêt à minimiser le coût de l’alimentation à un moment donné. Cette diminution est obtenue par des apports alimentaires inférieurs aux besoins des animaux. Les réserves corporelles (lipidiques, azotées, minérales, vitaminiques) constituées pendant la période de pâturage, permettent de faire face au déficit de l’apport. En principe, la reconstitution est réalisée sans problème lorsque l’herbe est abondante et que la capacité d’ingestion des animaux est suffisamment importante. Cette opération est économiquement rentable dans la mesure où l’alimentation au pâturage est de faible coût.
Animaux à besoins élevés pour lesquels entrent en jeu des impératifs physiologiques. Une telle situation se rencontre notamment en début de lactation, dans le cas des vaches laitières fortes productrices. Dans ce cas, la sous-alimentation (énergétique et minérale) des animaux est imposée, non pas par calcul stratégique, mais par capacité d’ingestion des animaux insuffisante face à des dépenses importantes.
EN RÉSUMÉ
Les activités de l’organisme animal sont à l’origine de dépenses ou besoins,
- d’une part en énergie,
- d’autre part en matériaux : glucose, acides aminés, éléments minéraux.
Ces dépenses couvrent l’entretien de l’animal, et éventuellement la réalisation de productions (croissance, engraissement, gestation, lactation, ponte, travail, etc).
L’objectif de l’alimentation rationnelle est de couvrir les besoins avec les apports alimentaires recommandés.
4.3 Besoins ou dépenses en énergie
La transformation des aliments au cours de la digestion apporte de l’énergie aux cellules de l’organisme. L’animal se procure l’énergie nécessaire à la couverture de ses besoins en utilisant les nutriments issus de la digestion des aliments et de l’absorption, ou en se servant de ses réserves corporelles en cas d’un jeûne ou d’une sous-alimentation.
Les dépenses énergétiques des animaux correspondent à une production de matières (fœtus, lait, tissus corporels, œuf, laine ou autre) contenant de l’énergie chimique, et une production de chaleur provenant du fonctionnement de l’organisme.
On peut distinguer 3 types de dépenses énergétiques :
la dépense énergétique d’entretien, liée au maintien de la vie et au fonctionnement normal de l’organisme dans les conditions d’élevage, sans tenir compte des productions,
les dépenses énergétiques de production, correspondant à l’énergie contenue dans les produits (fœtus, lait, tissus corporels, œuf, laine, ou autre),
et l’extra-chaleur qui est la dépense énergétique consécutive à l’utilisation des aliments pour satisfaire les dépenses d’entretien et de production. L’extra-chaleur se manifeste par une production de chaleur.
On peut écrire (équation 4.3) :
\[Dépense~totale = Dépense~d'entretien + Dépense~de~production + Extrachaleur\]
L’unité officielle de mesure de l’énergie est le joule (J) ou le kilojoule (kJ), mais on utilise aussi la calorie (cal) ou la kilocalorie (kcal). Le joule et la calorie sont liées par (équation 4.4) :
\[1~cal = 4,185~J~et~1~kcal = 4,185~kJ\]
4.3.1 La dépense en énergie pour l’entretien
La dépense d’entretien comprend l’énergie dépensée par le métabolisme de base et l’énergie nécessaire au maintien du poids de l’animal dans les conditions de vie normale.
4.3.1.1 Le metabolisme de base (Métabolisme basal)
C’est la dépense énergétique d’un animal à jeun, au repos, non endormi, placé à une température correspondant à la zone de neutralité thermique.
La zone de neutralité thermique est la plage de température où l’organisme n’effectue pas de dépense pour lutter contre l’excès de chaleur ou contre le froid (Tableau 4.1).
Tableau 4.1 Zone de neutralité thermique de différentes catégories d’animaux
| Catégorie d’animaux | Zone de neutralité thermique |
|---|---|
| Bovins | 5 à 20 °C |
| Chèvres | 10 à 20 °C |
| Veaux | 5 à 15 °C |
| Porcs de 40 à 80 kg | 20 à 23 °C |
| Porcs de 80 à 110 kg | 15 à l6 °C |
| Volailles | 15 à 25 °C |
| Poussins d’un jour | ≈ 32 °C |
| Poulettes de six semaines | ≈ 21 °C |
| Poules pondeuses | 15 à 20 °C |
| Lapins | 15 à 18 °C |
Le métabolisme de base augmente moins vite que le poids vif des animaux. Il n’est pas proportionnel au poids, mais plutôt à la surface corporelle et plus précisément au poids vif (kg) élevé à la puissance 0,75 qu’on appelle le poids métabolique (équation 4.5).
\[Poids~métabolique = (Poids~vif)^{0,75}\]
On exprime donc les dépenses du métabolisme de base par rapport au poids métabolique et non par rapport au poids vif.
La dépense du métabolisme de base est de l’ordre de \(70~kcal/kg~P^{0,75}\) pour l’homme et les espèces domestiques. Cette dépense varie avec :
l’espèce : \(75~kcal/kg~P^{0,75}\) chez les bovins, \(55~kcal/kg~P^{0,75}\) chez les ovins,
la race : elle est supérieure de 13 % chez les bovins laitiers comparés aux bovins à viande,
l’individu,
la saison,
la lumière
et la température.
4.3.1.2 Dépense énergétique d’entretien
Dans les conditions habituelles d’élevage, un animal n’est jamais placé dans les conditions définies pour la mesure du métabolisme de base. Il réalise une activité physique et vit dans des condifions climatiques parfois éloignées de celles du confort thermique.
La dépense énergétique d’un animal à l’entretien correspond à la production de chaleur de l’animal non productif à jeun. Elle comprend l’énergie dépensée par le métabolisme de base et l’énergie nécessaire au maintien de l’animal dans les conditions de vie normales (équation 4.6).
\[Entretien = Métabolisme~de~base + Thermorégulation + Activité~physique\]
4.3.2 La dépense en énergie pour la production
Les produits réalisés peuvent être exportés (lait, œufs, laine), ou stockés au sein de la masse corporelle (croissance).
Les quantités de lipides (Lip.), de protéines (Prot.) et de glucides (Glu.) fixées ou exportées permettent de calculer la quantité d’énergie contenue dans le produit réalisé, c’est-à-dire la dépense énergétique de production (DÉP) par (équation 4.7) :
\[DÉP~(kcal) = 5,5 * Prot. (g) + 9,4 * Lip. (g) + 4,1 * Glu. (g)\]
4.3.2.1 Dépense en énergie pour la production de lait
La dépense en énergie pour la production de lait dépend de la quantité de lait produite et de sa composition chimique. Le Tableau 4.2 présente les compositions moyennes d’un kg de lait de différentes espèces. La dépense occasionnée par la production de 1 kg de lait peut être calculée à partir de sa composition chimique.
Tableau 4.2 Composition moyenne d’un kg de lait de différentes espèces
| Matières grasses (g) | Protéines (g) | Lactose (g) | Énergie (kcal)* | |
|---|---|---|---|---|
| Lait standard de vache | 40 | 31 | 48 | 740 |
| Lait de chèvre | 32 | 30 | 43 | 670 |
| Lait de brebis | 63 | 56 | 44 | 1080 |
| Lait de truie (des 3 1ères semaines) | 69 | 46 | 55 | 1140 |
| Lait de lapine | 120 | 160 | 20 | 2250 |
(Verifiez les valeurs de l’énergie avec l’équation de la dépense énergétique !!!)
Lorsqu’il est difficile de connaitre la production laitière d’une femelle allaitante, on peut estimer la dépense en énergie pour la lactation à partir de la croissance de la portée. Le Tableau 4.3 donne des estimations de la production de lait de la brebis allaitante au cours des trois premières semaines de lactation à partir de la croissance entre 10 et 30 jours de la portée.
Tableau 4.3 Production laitière de la brebis (en litres) selon le GMQ 10-30j de sa portée.
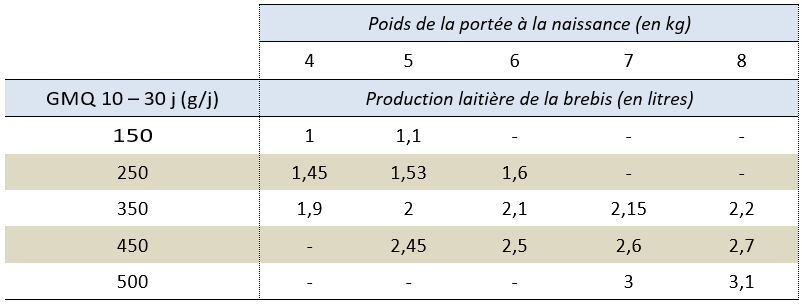
(S’essayer à estimer le besoin en énergie pour la production d’une brébis de chaque catégorie !!!).
4.3.2.2 Dépense en énergie pour la croissance
Elle correspond à l’énergie des protéines et des lipides fixés. La croissance est due essentiellement à une accumulation de protéines et de lipides corporels. On néglige l’énergie fixée sous forme de glucides, car les réserves glucidiques corporelles sont faibles.
La fixation énergétique s’effectue dans les muscles, les dépôts adipeux, les os, les viscères, la peau, le sang, etc. Elle dépend de la vitesse de croissance et de la composition chimique du croît.
La quantité d’énergie fixée est d’autant plus grande que le dépôt de gras est plus intense.
4.3.2.3 Dépense en énergie pour la gestation
La dépense en énergie pour la gestation correspond à la fixation d’énergie par le ou les fœtus, le placenta, les enveloppes, la paroi utérine et la glande mammaire. Elle devient importante au cours du dernier tiers \((1/3)\) de la gestation.
4.3.2.4 Dépense en énergie pour la production d’œufs
L’œuf entier de poule a la constitution moyenne suivante :
- coquille 9 %,
- albumen 61 %,
- vitellus 30 %.
La partie comestible d’un œuf de poule de \(60~g\) est composée en moyenne de :
- \(40~g\) d’eau ;
- \(6,7~g\) de protéines ;
- \(6,5~g\) de lipides ;
- \(0,2~g\) de glucides
- et \(0,5~g\) de minéraux.
- Il contient environ \(100~kcal\), soit \(1,6~kcal/g\) d’œuf.
4.3.2.5 Dépense en énergie pour la production de laine et de poils
Cette dépense est très faible. Elle est par exemple de :
- \(17~kcal/j\) pour la production annuelle de \(1~kg\) de laine mohair par une chèvre Angora,
- ou \(55~kcal/j\) pour la production annuelle d’une toison de \(4~kg\) par un mouton Mérinos.
4.3.2.6 Dépense en énergie pour la reconstitution des réserves corporelles
Chez les femelles en lactation, les réserves corporelles mobilisées participent à la couverture de la dépense énergétique lorsque l’apport énergétique est inférieur à la dépense. La reconstitution des réserves est nécessaire pour aborder un nouveau cycle de production. L’accroissement de la masse corporelle est surtout réalisé par une accumulation de lipides. \(1~kg\) de gain de masse corporelle correspond à une fixation énergétique de \(7500~kcal\) chez la vache laitière.
La masse corporelle est estimée imparfaitement à partir du poids vif car la masse du contenu digestif est très variable. L’appréciation et la notation de l’état corporel des animaux permettent d’estimer le niveau des réserves corporelles, ainsi que leurs variations dans le temps.
4.3.2.7 Dépense en énergie pour le travail musculaire (l’effort musculaire)
Tout travail musculaire provoque une augmentation de la consommation d’oxygène par rapport à celle enregistrée chez l’animal au repos. L’élévation de la consommation d’O2 reflète l’intensité du travail fourni et permet de quantifier la dépense calorifique consécutive au travail.
La consommation du cheval de selle de \(550~kg\), exprimée en litre d’O2/mn, s’accroît linéairement avec la vitesse de course, exprimée en m/mn. La relation est la suivante (équation 4.8) :
\[Quantité~d’O2~(en~litre/mn) = 3,78 + 0,097 * vitesse~(en~m/mn)\]
Ce même cheval au repos consomme \(1,65~litres~d'O_2/mn\). La dépense énergétique est calculée à partir de la consommation d’\(O_2\) pendant et après l’effort (exemple ultérieurement !). Après un effort, on enregistre une forte consommation appelée dette en \(O_2\), due au métabolisme du lactate produit par le muscle en état d’anaérobiose au moment de l’effort.
4.3.3 L’extra-chaleur
L’extra-chaleur ou la dépense d’extra-chaleur est consécutive à :
l’ingestion et la digestion des aliments, c’est-à-dire la prise des aliments, la mastication, le brassage, la progression du bol alimentaire dans le tube digestif, la chaleur des fermentations, l’activité glandulaire et métabolique des différentes parties du tube digestif. Il faut y ajouter la perte calorifique due à l’agitation des animaux avant la distribution des repas,
l’utilisation métabolique des nutriments absorbés. Par exemple, une mole de glucose fournit à la cellule \(473~kcal\) dans des conditions aérobies alors que la combustion complète d’une mole de glucose fournit \(673~kcal\). Toute l’énergie potentielle du glucose n’est pas utilisée par la cellule, il y a transformation incomplète de l’énergie avec un rendement de \(473/673~(≈ 0,7)\). Le complément, c’est-à-dire les 30 % restant de l’énergie, est perdu sous forme de chaleur.
Un animal qui ne réalise aucune production, couramment appelé animal à l’entretien (exemple : animal à poids constant, vache tarie non gestante à poids constant), a cependant deux types de dépenses :
une dépense énergétique d’entretien telle qu’elle a été précédemment définie
et une dépense d’extra-chaleur, dite extra-chaleur d’entretien.
Lorsque cet animal réalise une production, aux dépenses précédentes s’ajoutent (Figure 4.2):
- une dépense énergétique de production et
- une dépense d’extra-chaleur appelée extra-chaleur de production.
On peut donc écrire (équation 4.8) :
\[Extrachaleur~totale = Extrachaleur~d'entretien + Extrachaleur~de~production\]
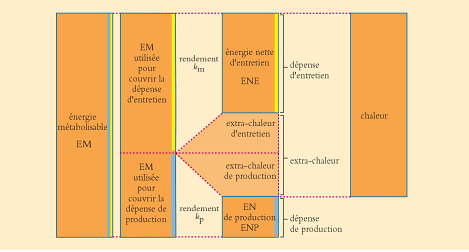
Figure 4.2 Utilisation de l’énergie métabolisable
On définit également l’extra-chaleur comme correspondant à l’augmentation de production calorique due au fait qu’un animal passe de l’état de jeûne à l’état d’animal alimenté (équation 4.9) :
\[ECt = Calimenté~–~Cjeun\] avec :
- \(ECt = Extrachaleur~totale\),
- \(Calimenté = production~de~haleur~de~l’animal~alimenté\),
- \(Cjeun = production~de~chaleur~de~l’animal~à~jeun\).
La Figure 4.3 fait apparaître les différentes origines de la chaleur produite : entretien, extra-chaleur d’entretien et extra-chaleur de production.
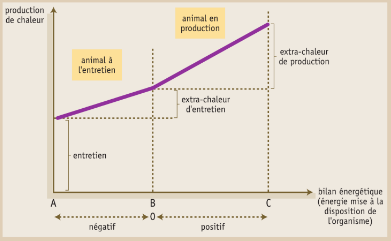
Figure 4.3 Évolution de la production de chaleur en fonction du bilan énergétique.
- Le point A correspond à un animal à jeun et à poids constant, qui produit cependant de la chaleur en mobilisant des reserves corporelles pour couvrir ses dépenses d’entretien (cas théorique).
- Le point B correspond à un animal à l’entretien et alimenté,
- et le point C, à un animal en production et alimenté.
En résumé
Les dépenses énergétiques des animaux sont de 3 types :
- la dépense d’entretien qui comprend l’énergie dépensée par le métabolisme de base et l’énergie nécessaire au maintien de l’animal dans des conditions de vie normale :
\[Entretien = métabolisme~de~base + thermorégulation + activité~physique\]
Le métabolisme de base est la dépense énergétique d’un animal à jeun, au repos, placé à une température correspondant à la zone de neutralité thermique,
la dépense de production correspond à l’énergie contenue dans les produits réalisés : lait, gain de poids, œufs, ou autre,
la dépense liée à l’utilisation des aliments, l’extra-chaleur, consécutive à l’ingestion et à la digestion des aliments, et aux transformations chimiques des nutriments mis à la disposition de l’organisme pour couvrir les dépenses.
4.3.4 Les différentes étapes de l’utilisation de l’énergie
L’utilisation des aliments par les animaux s’accompagne de nombreuses transformations dans l’appareil digestif puis dans les tissus. Les composants organiques (les glucides, les proteines et les lipides) sont dégradés progressivement, chaque étape entraînant des pertes organiques et calorifiques (énergétiques) qui varient selon les aliments et selon les animaux.
4.3.4.1 L’énergie brute
L’énergie brute (EB) est la quantité d’énergie contenue dans l’aliment. Elle est déterminée par combustion d’un échantillon dans une bombe calorimétrique, ou estimée par des formules à partir de la composition chimique de l’aliment. La quantité d’énergie brute varie selon la nature de l’aliment en fonction des nutriments présents dans celui-ci. L’EB n’est jamais valorisée complètement par l’animal (Figure 4.4).
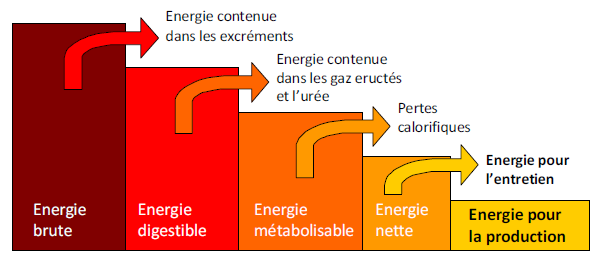
Figure 4.4 Les différentes étapes de l’utilisation de l’énergie des aliments
En effet, selon la digestibilité de la ration, une fraction plus ou moins importante de l’EB se retrouve dans les matières fécales et est donc perdue. L’énergie résiduelle s’appelle l’énergie digestible (ED). Une fraction de l’ED est ensuite perdue via les urines et les gaz, l’énergie restante s’appelant l’énergie métabolisable (EM). Au niveau cellulaire, l’EM est en partie dissipée sous forme d’extrachaleur (de façon simplifiée, surplus de production de chaleur chez l’animal qui a fait un repas), le solde étant l’énergie nette (EN) qui est disponible pour les cellules animales.
4.3.4.2 L’énergie digestible
L’énergie digestible (ED) est obtenue par différence entre l’énergie brute EB et l’énergie contenue dans les matières organiques fécales (EF) (équation 4.10) :
\[ED = EB - EF\]
La digestibilité de l’énergie (dE) est le rapport entre l’ED et l’EB \(dE = ED/EB\) d’où (équation 4.11) :
\[ED = EB * dE\]
Pour un type d’animal donné, la teneur de l’aliment en parois est le principal facteur de variation de la digestibilité de la matière organique et donc du coefficient \(dE\). Il en résulte une grande variabilité de ce coefficient entre les aliments.
4.3.4.3 L’énergie métabolisable et ses facteurs de variation
L’énergie métabolisable (EM) est égale à l’énergie digestible diminuée de l’énergie contenue dans les gaz combustibles, surtout le méthane, issus des fermentations digestives (EG) et de l’énergie contenue dans les urines (EU) (équation 4.12) :
\[EM = ED - EG - EU~~ou~~EM = EB - (EF + EG + EU)\]
On peut aussi écrire, en faisant apparaître le rapport \(\frac{EM}{ED}\), que \(EM = ED*\frac{EM}{ED}\).
L’énergie métabolisable correspond à la quantité d’énergie alimentaire utilisable par les tissus de l’organisme. Elle est sensiblement constante pour un aliment quelle que soit la fonction (croissance, lactation, engraissement) pour laquelle elle est utilisée. L’EM sert à faire face aux dépenses énergétiques liées à l’entretien et à la production (équation 4.13).
\[EM_{totale} = EM_{entretien} + EM_{production}\]
Facteurs de variation de l’EM
L’importance de la production de gaz témoigne de l’activité de la flore digestive. L’énergie contenue dans le méthane rejeté peut atteindre :
- 8 % de l’EB des fourrages distribués à des ruminants ;
- 3 à 4 % de l’EB des foins ingérés par le cheval ;
- 1 % de l’EB ingérée par le porc.
Pour rappel, les nutriments absorbés ne sont pas totalement dégradés dans l’organisme et une partie est excrétée dans l’urine, en particulier sous forme d’urée ou d’acide urique. L’énergie urinaire correspond à 3 à 5 % de l’EB, et son importance varie avec :
l’animal : les animaux fixant une grande quantité de protéines excrètent peu de matières azotées urinaires, et perdent donc peu d’énergie dans leurs urines ;
l’aliment : les pertes urinaires sont accrues si les apports azotés sont supérieurs aux besoins des animaux, ou sont excessifs par rapport au contenu énergétique de la ration, ou sont mal équilibrés en acides aminés.
4.3.4.4 L’énergie nette et ses facteurs de variation
Une partie seulement de l’EM contribue à couvrir les dépenses d’entretien et de production : c’est l’énergie nette. L’autre partie est dissipée sous forme de chaleur appelée l’extra-chaleur (équation 4.14) :
\[EN = EM - extrachaleur~~ou~~EN = k * EM\]
où k est le rendement global de la transformation de l’EM en EN : \(k = \frac{EN}{EM}\).
On peut déterminer l’EN à partir de l’EB des aliments en tenant compte de l’ensemble des pertes énergétiques facilement mesurables (fèces, urines et gaz) et des pertes calorifiques d’extra-chaleur (équation 4.15) :
\[EN = EB - (EF + EU + EG) - extrachaleur\]
Ces différentes pertes sont très variables selon la fonction physiologique réalisée par l’animal et la composition des aliments. On est conduit à caractériser chaque fonction par son rendement \(k = \frac{EN}{EM}\) ; par exemple (équation 4.16) :
\[k_{m}~(maintenance)~pour~l'entretien = \frac{EN~utilisée~pour~l'entretien}{EM~utilisée~pour~l'entretien}\]
De même, on peut définir (équation 4.17) :
\[k_{L}~(lactation)~pour~la~production~de~lait = \frac{EN~utilisée~pour~la~production~de~lait}{EM~utilisée~pour~la~production~de~lait}\]
équation 4.18:
\[k_{f}~(fattening)~pour~l'engraissement = \frac{EN~utilisée~pour~l'engraissement}{EM~utilisée~pour~l'engraissement}\]
etc.
Facteurs de variation du rendement k
Il y a deux facteurs de variation qui jouent un rôle important : - la demande en nutriments qui dépend de la fonction physiologique, et - l’offre de nutriments liée aux caractérstiques de la ration.
(Pouvez-vous, en vous servant de k, expliquer ces variations possibles ???)
EN RÉSUMÉ
La valeur énergétique d’un aliment peut être estimée :
en énergie brute (EB), déterminée par combustion de l’aliment ou à partir de sa composition chimique,
en énergie digestible (ED), égale à l’énergie brute diminuée de l’énergie des fèces \((ED = EB - EF)\),
en énergie métabolisable (EM), égale à l’énergie digestible diminuée de l’énergie des gaz et de l’urine \((EM = ED - EG - EU)\),
en énergie nette (EN), égale à l’énergie métabolisable diminuée de l’extra-chaleur (EC) \((EN = EM - EC)\).
L’énergie nette est utilisée pour couvrir les dépenses d’entretien et de production. Le rendement \(k = \frac{EN}{EM}\) est le rendement de la transformation de l’énergie métabolisable en énergie nette. Il dépend des caractéristiques de l’aliment et de la fonction considérée. Différents coefficients sont présentés à la Figure 4.5.
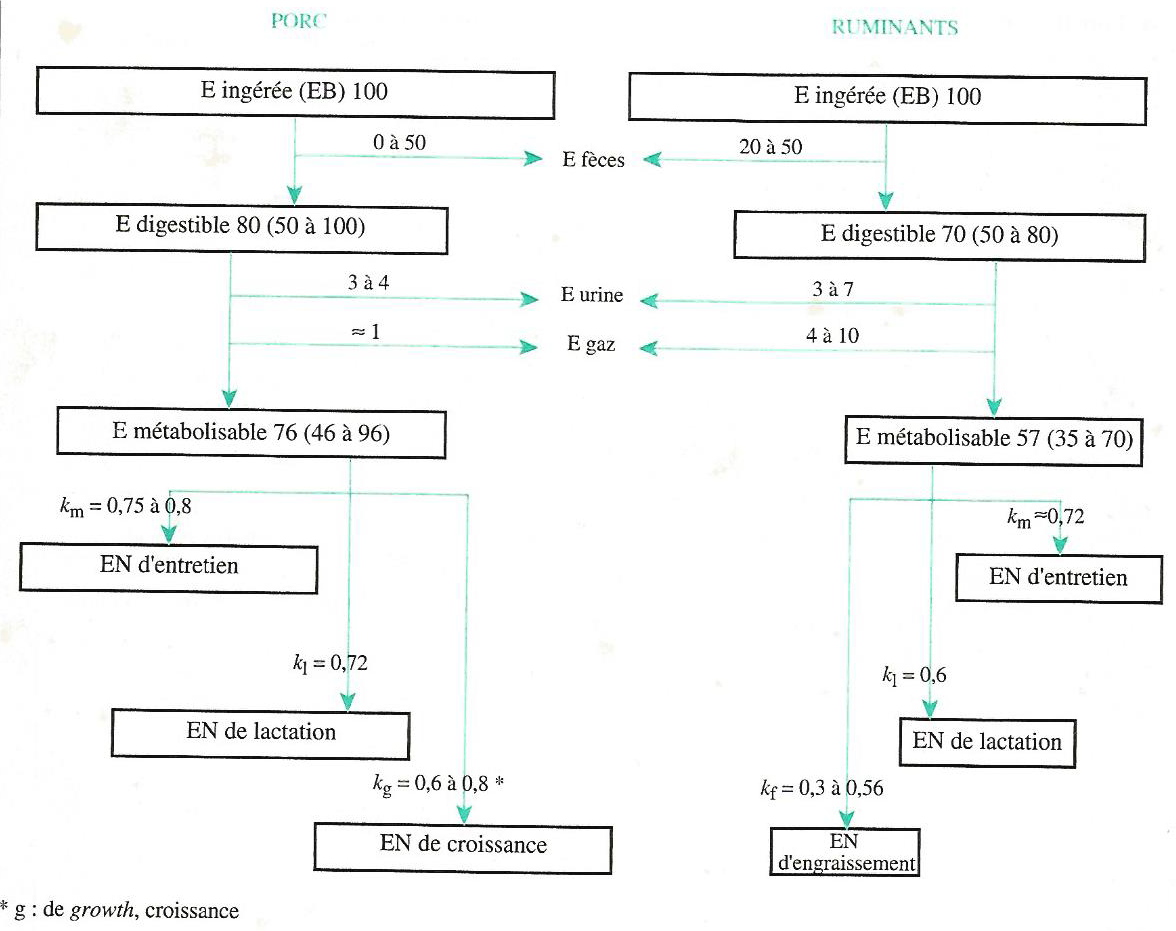
Figure 4.5 Schéma général de l’utilisation de l’énergie chez le porc et le ruminant
4.3.5 Retour sur les systèmes d’expression des besoins et apports en énergie
Après les détails précédents sur les étapes de l’utilisation de l’énergie, ce retour sur les systèmes d’expression peut être mieux abordé.
4.3.5.2 Système utilisant l’énergie digestible ED (lapins et porcs)
Les mesures de la digestibilité sont simples à réaliser chez les animaux soumis à ce système (lapins et porcs). On obtient la valeur énergétique des aliments en multipliant l’énergie brute de l’aliment par la digestibilité (ou coefficient de digestibilité) de l’énergie : \(ED = EB * dE\).
Les besoins sont quant à eux, déterminés à partir de 3 éléments :
- la dépense, exprimée en kcal d’EN,
- le rendement k, et
- le rapport \(\frac{EM}{ED}\).
On adopte pour k et \(\frac{EM}{ED}\) des valeurs moyennes (équation 4.19) :
\[Besoin~en~ED = dépense * \frac{1}{k} * \frac{1}{(EM/ED)}\]
Par exemple, \(1~kg\) de lait de truie contient \(1140~kcal\), le rendement \(k_{l}\) est de \(0,72\), et le rapport \(\frac{EM}{ED}\) est en moyenne de \(0,96\) chez les porcs. La valeur énergétique de ce kg de lait engendre un besoin de \(\frac{1140}{0,72 * 0,96} = 1649~kcal~d’ED\) à apporter par l’aliment.
On fait remarquer que le système utilisant l’ED surestime la valeur énergétique des protéines, car il ne tient pas compte des pertes urinaires dues à l’oxydation incomplète des acides aminés par l’animal : >> 1 g d’acide aminé digestible a une valeur de 5,5 à 5,6 kcal d’ED, alors que, après catabolisme, sa valeur n’est plus que de 4,4 à 4,6 kcal d’EM. La différence correspond à l’énergie retrouvée dans les urines.
4.3.5.3 Système utilisant l’énergie métabolisable EM
Le système fondé sur l’énergie métabolisable est utilisé pour les volailles, car les urines et les matières fécales sont mélangées dans les fientes au niveau du cloaque. De ce fait, on ne peut pas raisonner facilement en énergie digestible pour ces animaux. Il existe des équations de prévision des besoins totaux journaliers. Pour déterminer la valeur énergétique des aliments, on peut appliquer la formule générale \(EM = EB * dE * \frac{EM}{ED}\), mais on dispose de relations particulières à chaque aliment.
Ce système est également appliqué aux veaux de boucherie chez lesquels le rendement global \(k\) est assez constant (\(k = 0,69\)). Chez ces animaux, l’étape de l’énergie nette n’apporte pas de précision complémentaire. Ce système pourrait être appliqué aux porcs.
Les besoins peuvent être déterminés à partir de la dépense et du rendement \(k\) : \(Besoin~en~EM = \frac{dépense}{k}\). Par exemple, le besoin journalier de croissance du veau est calculé par la formule : \(Besoin~en~EM = \frac{contenu~énergétique~ducroît}{0,69}\).
Comparée à l’énergie digestible, l’énergie métabolisable conduit à une meilleure estimation de la valeur énergétique des aliments riches en protéines.
4.3.5.4 Système utilisant l’énergie nette EN (pour les ruminants)
Variation des rendements d’utilisation de l’EM (k)
La Figure 4.6 présente les variations des rendements \(k_{m}\) (entretien), \(k_{l}\) (lactation) et \(k_{f}\) (engraissement) en fonction du rapport \(q = \frac{EM}{EB}\) caractérisant la qualité de l’aliment. Ce rapport qui est la concentration en EM de l’aliment, est utilisé parce qu’il évolue très parallèlement à la digestibilité de l’énergie.
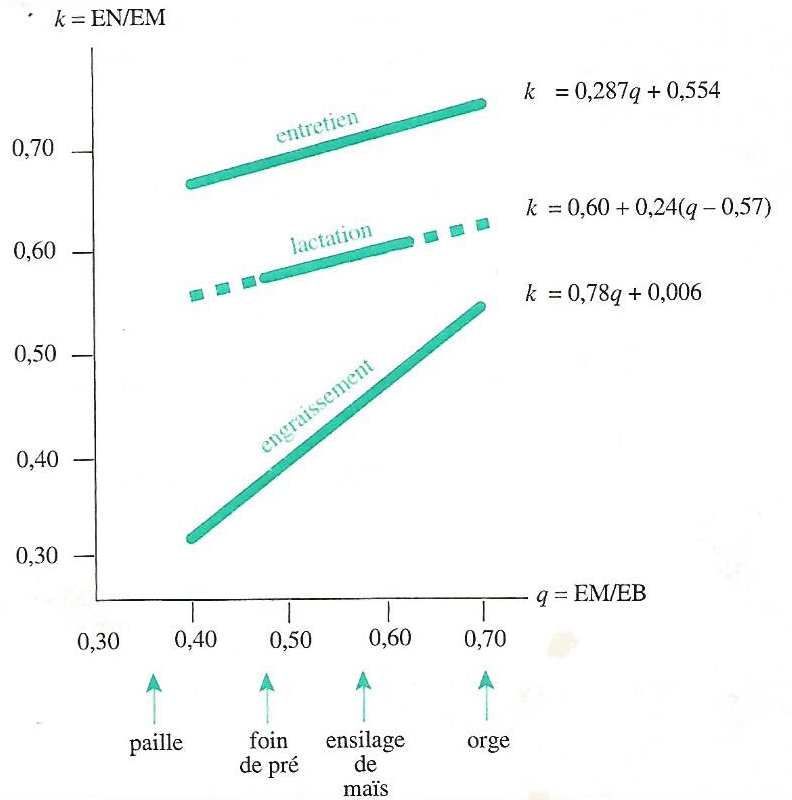
Figure 4.6 Variations des rendements \(k_{m}\), \(k_{l}\) et \(k_{f}\) chez les ruminants en fonction de la concentration en EM de la ration, (\(q = \frac{EM}{EB}\)).
On constate que :
- quel que soit l’aliment, \(k_{m}\) > \(k_{l}\) > \(k_{f}\),
- les deux droites correspondant à \(k_{m}\) et \(k_{l}\) sont sensiblement parallèles, il existe donc un rapport constant entre \(k_{m}\) et \(k_{l}\) de \(\frac{k_{m}}{k_{l}} ≈ 1,2\) pour tous les aliments,
- le rendement \(k_{f}\) varie très fortement selon les aliments et il n’y a pas de rapport constant entre \(k_{f}\) et \(k_{m}\), ni entre \(k_{f}\) et \(k_{l}\).
Compte tenu donc des différences de rendements d’utilisation de l’EM selon les fonctions physiologiques, on peut définir au moins trois valeurs énergétiques nettes pour chaque aliment :
\[EN~d'entretien = EM * k_{m}\]
\[EN~de~lactation = EM * k_{l}\]
\[EN~d'engraissement = EM * k_{f}\]
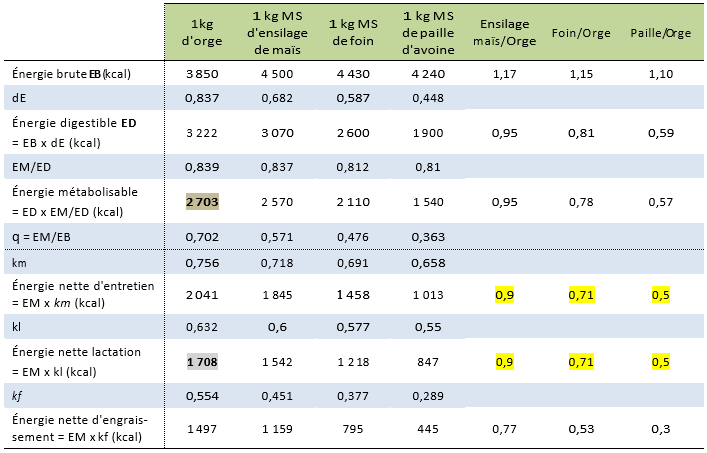
Les exemples du Tableau 4.4 montrent que :
- la fonction physiologique considérée a une influence sur la valeur en énergie nette de l’aliment ;
- pour une même fonction, il existe des écarts de valeur en énergie nette entre aliments, écarts plus importants pour l’engraissement que pour l’entretien ou la lactation ;
- la valeur énergétique d’un aliment exprimée par rapport à l’orge est la même pour l’entretien et la lactation. Pour exemple, pour le foin, elle est égale à 0,71.
Tableau 4.4 Valeurs énergétiques de quelques aliments chez les ruminants (Gadoud et al., 1992).
Le Système de l’Unité Fourragère (UF) : définition et principe général
On rappelle qu’en pratique, deux fonctions différentes au moins sont réalisées par un même animal avec une importance relative très variable. Pour simplifier et éviter de multiplier le nombre de valeurs énergétiques d’un aliment, le système français (INRA, 1978, modifié en 1988) a retenu deux situations :
la combinaison entretien + lactation (système « lait ») : compte tenu du rapport constant \(\frac{k_{m}}{k_{l}} ≈ 1,2\), les aliments ont, en valeur relative, la même valeur énergétique nette pour l’entretien et la lactation ;
une combinaison entretien + engraissement (système «viande») correspondant à des animaux à croissance intensive utilisant un tiers \((1/3)\) de l’énergie nette pour la production et deux tiers \((2/3)\) pour l’entretien.
Concrètement :
- le système « Lait » est utilisé pour les femelles en lactation, celles gestantes ou taries, les femelles d’élevage, les animaux à l’entretien et les animaux réalisant une croissance modérée ;
- le système « Viande » est utilisé pour les bovins ayant un GMQ supérieur à 1000 g/j et pour les agneaux et chevreaux à croissance élevée.
De plus, le mode d’expression de la valeur énergétique des aliments par rapport à celle de l’orge, dans ce système, évite de parler de kcal ou d’autres types d’unités d’énergie, mais se réfère à la valeur énergétique d’un aliment de référence, l’orge, appelée unité fourragère (UF).
Pour une brève histoire, la notion d’unité fourragère a été introduite en Scandinavie au début du 20è siècle. Elle a été reprise en France vers 1950 et est toujours utilisée dans le système français actuel d’estimation des besoins et d’expression de la valeur énergétique des aliments, proposés par l’INRA.
L’unité fourragère est la valeur en énergie nette d’un kg d’orge de référence. On définit ainsi la valeur énergétique (VE) d’un aliment en UF comme étant le rapport :
\[VE_{aliment}~(UF) = \frac{énergie~nette~de~l'aliment~(kcal)}{énergie~nette~de~1~kg~d'orge~de~référence~(kcal)}\]
Compte tenu des 2 systèmes retenus (Lait et Viande), on a donc 2 unités fourragères correspondant aux valeurs énergétiques nettes de l’orge de référence :
- dans le système Lait, Unité Fourragère « Lait » (UFL) ;
- dans le système Viande, Unité Fourragère « Viande » (UFV).
Chaque aliment a ainsi deux valeurs énergétiques rapportées à celles de l’orge de référence et exprimées en UFL et en UFV. Les apports alimentaires recommandés sont exprimés en UFL ou en UFV, suivant le type d’animal considéré (production de lait, ou production de viande). Il existe également une unité fourragère cheval, UFC (pas développée !!!).

(Je devrais reproduire ou indiquer le tableau 5.2 référé !)
Le système de l’Unité Fourragère Lait UFL
L’UFL est la valeur énergétique nette de lactation d’un kg d’orge de référence distribué à une femelle laitière dont on a couvert la dépense d’entretien. Cette énergie est exportée dans le lait. Elle correspond à \(1708~kcal~d'ENL\) (Tableau 4.4), valeur arrondie à \(1700~kcal~d'ENL\).
\[1~UFL = 1700~kcal~d'ENL\]
Un exemple de calculs est présenté à la la capture de page ci-haut.
Le système de l’Unité Fourragère Viande UFV
a. La notion de rendement global d’utilisation de l’EM k_{mf}
Il n’existe pas de rapport constant entre \(k_{m}\) et \(k_{f}\). Pour l’animal à croissance élevée, on définit un rendement global \(k_{mf}\) d’utilisation de l’énergie métabolisable des aliments pour l’entretien et l’engraissement (Figure 4.7).
\[k_{mf} = \frac{EN~d'~Entretien~+~EN~Viande}{EM~Totale} = \frac{ENE+ENV}{EMT} = \frac{ENEV}{EMT}\]
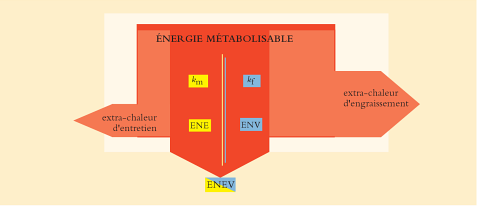
Figure 4.7 Utilisation de l’EM pour l’entretien et l’engraissement
Le rendement global \(k_{mf}\) est calculé à partir :
- des rendements partiels \(k_{m}\) et \(k_{f}\), et
- de l’importance relative de l’EN d’entretien (ENE) et de l’EN de production de viande (ENV*), c’est-à-dire du niveau de production NP :
\[NP = \frac{ENE+ENV}{ENE}\]
On aboutit à :
\[k_{mf} = \frac{k_{m}*k_{f}*NP}{k_{f}+(NP-1)*k_{m}}\]
b. Le choix d’un Niveau de Production (NP)
Le rendement \(k_{mf}\) calculé pour un aliment dépend du NP, donc de l’importance relative de l’entretien et de la croissance. Il peut donc prendre une infinité de valeurs. Pour éviter cet inconvénient, il est nécessaire de fixer un seul niveau de production pour les animaux destinés à la production de viande.
Sur la base du système français, on classe les bovins en croissance et à l’engrais en deux catégories :
des animaux ayant un NP compris entre 1 et 1,4 avec une moyenne de 1,2 : animaux au voisinage de l’entretien, sujets réalisant une croissance modérée comme les femelles d’élevage (GMQ < 1000 g pour les bovins en général). A ce stade, les dépenses d’entretien étant nettement prépondérantes, on utilise le système des UFL ;
des animaux ayant un \(NP > 1,4\) avec une moyenne de 1,5 : animaux en croissance intensive (GMQ > 1000 g pour les bovins) ou à l’engrais. A ce stade, les dépenses d’entretien sont plus faibles. La valeur \(NP = 1,5\) est donc choisie pour ces animaux. Dans ce cas, les deux tiers de l’EN sont utilisés pour l’entretien et un tiers pour la production : \(NP = \frac{2+1}{2} = 1,5\). (Voir formule de NP pour comprendre !).
On définit ainsi l’UFV comme la quantité d’énergie nette fournie par \(1~kg\) d’orge de référence pour l’entretien et l’engraissement à un animal en croissance ayant un Niveau de Production NP de 1,5.
L’EM de l’orge (2703 kcal – Tableau 4.4) est utilisée pour l’entretien et la croissance avec le rendement :
\[k_{mf} = \frac{0,756 * 0,554 * 1,5}{0,554 + (1,5-1) * 0,756} = 0,674\]
\(1~kg\) d’orge fournit donc \(2703 * 0,674 = 1822~kcal~d'ENEV\), valeur arrondie à \(1820~kcal\).
Bien que \(k_{f}\) soit inférieur à \(k_{l}\), la valeur de l’UFV (\(1820~kcal\)) est supérieure à celle de l’UFL (\(1700~kcal\)), car \(k_{mf}\) est supérieur à \(k_{l}\) pour l’orge. Des exemples de calculs sont présentés dans la capture de page ci-dessous.

EN RÉSUMÉ
C’est l’étape ultime de la transformation d’un aliment par l’animal qui constitue le niveau le plus pertinent pour estimer la valeur énergétique de cet aliment. Il est donc toujours souhaitable de pouvoir exprimer les besoins des animaux et les apports des aliments en énergie nette ; c’est le cas des ruminants et du cheval.
Cependant, les variations du rendement \(k = \frac{EN}{EM}\) selon la fonction physiologique considérée (entretien, lactation, engraissement, etc.) ne permettent pas dans tous les cas d’utiliser l’énergie nette. C’est pourquoi on utilise l’énergie métabolisable chez les volailles et les veaux de boucherie, et l’énergie digestible chez les porcs et les lapins.
Chez les ruminants, le système Unité Fourragère distingue deux situations types :
le système « lait », utilisé pour les femelles en lactation, les femelles gestantes ou taries, les femelles d’élevage, les animaux à l’entretien et les animaux à croissance modérée ;
le système « viande », utilisé pour les animaux à croissance élevée, en particulier les bovins dont le GMQ est supérieur à \(1000~g/j\).
L’unité retenue, appelée unité fourragère (UF), correspond à l’énergie nette d’un kg d’orge de référence. On distingue : l’UF lait (UFL) et l’UF viande (UFV). Chaque aliment a deux valeurs énergétiques, exprimées en UFL et UFV, et les apports alimentaires recommandés sont donnés en UFL ou en UFV selon le type d’animal.
4.4 Expression des besoins et des apports en matières azotées
Les matières azotées sont des substances plastiques dont le rôle principal est l’élaboration des protéines de constitution de l’organisme, des productions ou de protéines fonctionnelles (enzymes, hormones, anticorps, etc.). Leur second rôle est de fournir de l’énergie, lorsque les AA absorbés dépassent les possibilités de synthèse de l’organisme (trop d’azote par rapport à l’énergie ingérée). Ces AA excédentaires sont dégradés pour fournir de l’énergie. Ce qui s’accompagne de production d’azote éliminée dans les urines sous forme d’urée.
4.4.1 Les dépenses azotées
On considère deux types de dépenses :
- Le renouvellement constant des tissus de l’organisme et les synthèses de substances liées à son fonctionnement : il s’agit des dépenses d’entretien, et
- L’exportation des protéines des produits : gain de poids, fœtus, lait, œufs, etc.
L’alimentation azotée des animaux repose sur des données fondamentales :
les acides amines (AA) constituent la seule forme azotée utilisable par les cellules animales,
les cellules ont des besoins en acides aminés indispensables (AAI) et les autres AA,
les synthèses nécessitent de l’énergie : il y a une interdépendance entre l’alimentation énergétique et l’alimentation azotée.
La carence en azote se traduit par une perte d’appétit et donc une diminution des performances. A l’inverse, un excès engendre des disfonctionnements de l’organisme :
- modification du faciès microbien,
- fatigue hépatique ou rénale,
- alcalose sanguine, etc.
En outre, un déséquilibre azoté augmente les pertes d’azote (polluant pour l’environnement) dans les urines et les matières fécales.
4.4.2 Les dépenses azotées d’entretien
Les cellules de l’organisme sont en perpétuel renouvellement et subissent donc des pertes obligatoires de substances azotées. L’urine et les fecès sont les deux voies pricipales d’élimination des constituants qui se dégradent et leur importance relative varie avec l’espèce animale :
l’urine est la voie principale d’élimination des déchets azotés chez les monogastriques (90 % chez l’homme, 80 % chez le porc, 60 % chez les oiseaux),
la voie fécale prédomine chez les ruminants (70 à 80 %).
4.4.3 Les dépenses azotées de production
Pour les dépenses azotées de production, on évalue l’importance globale (la quantité) et la composition en acides aminés (l’aspect qualitatif).
4.4.3.1 La dépense azotée pour la croissance
Le dépôt azoté quotidien correspond aux dépôts dans les différents compartiments corporels (muscles, os, graisse, peau, sang, etc.). Son importance dépend :
- de la croissance, elle-même liée à l’âge, au type génétique et au sexe,
- du mode d’alimentation : à volonté ou restreint,
- des conditions d’élevage et de production, poids et âge d’abattage.
4.4.3.2 La dépense azotée pour la gestation
Cette dépense azotée correspond aux matières azotées fixées dans l’utérus, et par le ou les fœtus et les annexes. On ne tient compte d’un besoin azoté spécifique qu’au cours du dernier tiers (\(1/3\)) de la gestation.
4.4.3.3 La dépense azotée pour la lactation
La dépense azotée pour la lactation dépend de la composition du lait et est donc variable selon les espèces. Mais dans une espèce donnée, la dépense azotée varie avec :
- la race : le taux azoté varie selon les types génétiques de \(30~à~35g/kg\),
- le stade de lactation : au cours de la lactation, les teneurs en matières grasses et matières azotées évoluent en sens inverse de la quantité de lait produite (phénomène de dilution).
4.4.3.4 La dépense azotée pour la production d’œufs
Les œufs sont riches en matières azotées (environ 12 % de leur poids). La ponte d’un œuf de \(60~g\) représente donc une dépense de \(7~g\) de matières azotées, soit 3,5 fois les dépenses azotées d’entretien d’une poule de 2,5 kilos (Gadoud et al., 1992).
4.4.3.5 La dépense azotée pour la production de laine
La laine est un tissu de composition complexe où prédomine la kératine (protéine spécifique des phanères) riche en acides aminés soufrés. Ces derniers peuvent en cas de carence devenir des facteurs limitants de la pousse de la laine ou des poils chez les animaux à fourrure.
4.4.3.6 La dépense azotée pour le travail musculaire
Les dépenses azotées liées au travail musculaire ne sont pas connues avec précision. Il est admis qu’elles augmentent moins vite que les dépenses en énergie. Cependant, elles peuvent augmenter plus rapidement si les apports énergie sont insuffisants dans le cas d’efforts très intenses.
4.4.4 La couverture des dépenses azotées
Dans toutes les espèces, la synthèse cellulaire des protéines suppose la présence simultanée de 22 acides aminés que sont : le glycocolle, l’alanine, la sérine, la thréonine, la valine, la leucine, l’isoleucine, l’acide aspartique, l’acide glutamique, l’acide hydroxyglutamique, l’arginine, la lysine, la cystine, la méthionine, la phénylalanine, la tyrosine, le trypophane, l’histidine, la proline, l’hydroxyproline, N° 21 = la sélénocystéine découverte en 1999, et N° 22 = la pyrrolidine découverte en 2002.
Ils se répartissent en deux grandes catégories selon les possibilités de synthèse par l’organisme
4.4.4.1 Les acides aminés essentiels
Les acides aminés essentiels ou indispensables sont au nombre de 10. Certains d’entre eux sont dits indispensables au sens strict : l’animal est incapable de les synthétiser. Ce sont la lysine et la thréonine pour toutes les espèces. Et d’autres essentiels au sens large : ils peuvent être synthétisés à partir d’autres acides aminés. Mais cette synthèse est trop faible pour assurer les besoins de l’animal. Il s’agit de l’arginine, la méthionine, l’histidine, la phénylalanine, l’isoleucine, le tryptophane, la leucine et la valine.
4.4.4.2 Les acides aminés banals
Ce sont les autres acides aminés. L’animal peut en synthétiser en quantité et vitesse suffisante. Cependant, leur apport par l’aliment ne doit pas être négligé car ils économisent les acides aminés essentiels.
Si un acide aminé essentiel vient à manquer au niveau de la protéosynthèse, elle diminue et peut s’arrêter. Les performances chutent et l’indice de consommation augmente. L’excrétion urinaire d’azote augmente puisque les autres acides aminés présents se retrouvent en excès. Cet acide aminé, dont la quantité insuffisante limite la protéosynthèse, est appelé facteur limitant : son apport règle le niveau de la protéosynthèse et donc l’efficacité de l’utilisation des protéines de l’aliment (Figure 4.8).
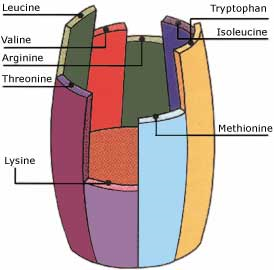
Figure 4.8 Notion de facteur limitant (la lysine), adaptataion du baril de Liebig
4.4.5 Utilisation des matières azotées alimentaires
Pour rappel, les matières azotées alimentaires ne sont pas toutes utilisées par l’animal. Une partie est rejetée dans les déjections (urine, fecès).
L’utilisation digestive de l’azote des aliments conduits à l’élimination fécale de la fraction non digérée. L’importance relative de cette fraction dépend de l’espèce animale et des aliments constituant la ration.
L’utilisation métabolique des acides aminés est la fraction des acides aminés sanguins fixés pour la synthèse des protéines corporelles. L’excrétion d’azote urinaire en est une conséquence. L’importance de cette fraction dépend de l’adéquation en quantité (apport de matières azotées totales) et en qualité (apport d’acides aminés) de l’apport azoté, aux besoins de l’animal (Figure 4.9).
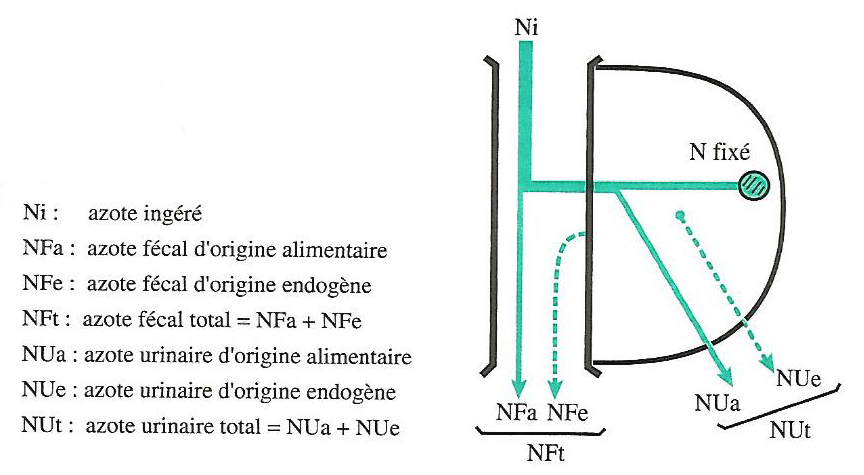
Figure 4.9 Utilisations digestive et métabolique de l’azote alimentaire
L’importance des pertes d’azote peut ainsi être caractérisée : 1. au niveau digestif par (cf. chapitre digestion) :
- le Coefficient d’Utilisation Digestive apparente (CUDa) et,
- le Coefficient d’Utilisation Digestive réel (CUDr)
- au niveau métabolique par :
- le coefficient d’utilisation métabolique apparent
\[CUM_{apparent} = \frac{N~fixé~apparemment}{N~absorbé~apparemment} = \frac{N_{i}-(NF_{t}+NU_{t})}{N_{i}-NF_{t}}\]
- et le coefficient d’utilisation métabolique réel, appelé encore valeur biologique (VB) :
\[CUM_{réel} = \frac{N~fixé~réellement}{N~absorbé~réellement} = \frac{N_{i}-(NF_{a}+NU_{a})}{N_{i}-NF_{a}}\]
Ou encore :
\[CUM_{réel} = \frac{N_{i} - [(NF_{t} - NF_{e})+(NU_{t} - NU_{e})]}{N_{i} -(NF_{t} - NF_{e})}\]
Ainsi, quelles que soient l’espèce animale concernée, pour améliorer l’efficacité de l’apport azoté alimentaire, il faut :
- réduire la fraction de l’azote indigestible rejetée dans les fèces ;
- réduire la fraction excrétée dans l’urine, c’est-à-dire bien ajuster la quantité des apports aux besoins des animaux et rechercher un équilibre optimal des protéines du régime en acides aminés, en particulier en acides aminés essentiels.
EN RÉSUMÉ
Comme les dépenses en énergie présentées plus haut, les dépenses en matières azotées distinguent des dépenses pour l’entretien et des dépenses pour la production. La dépense azotée d’entretien correspond au renouvellement perpétuel des cellules. Les dépenses de production (croissance, gestation, lactation, ponte, …) dépendent de l’importance des productions et de la teneur en azote des produits.
Grâce à des réactions de transamination, certains acides aminés sont synthétisés dans l’organisme à partir d’autres acides aminés : ce sont les acides aminés banals.
Pour d’autres, ces réactions n’existent pas ou sont insuffisantes : ce sont les acides aminés essentiels ou indispensables : lysine, méthionine, thréonine, tryptophane, …
Pour que la protéosynthèse puisse s’effectuer, il faut fournir aux cellules tous les acides aminés nécessaires, et dans une proportion définie par la composition de la protéine à synthétiser.
Pour obtenir une efficacité maximale de l’apport azoté alimentaire, il faut : - un taux azoté suffisant ; - un équilibre optimal des protéines du régime en acides aminés indispensables.
4.4.6 Retour sur l’expression des apports et des besoins en matières azotées
Pour rappel, le système d’expression doit exprimer les apports et les besoins dans une même unité. On peut exprimer les besoins à plusieurs niveaux (Figure 4.10) :
- au niveau de l’apport des matières azotées alimentaires (au niveau de l’aliment) ;
- au niveau de l’absorption intestinale ;
- au niveau cellulaire où a lieu la dépense, et où cette dépense peut être évaluée en protéines et/ou en acides aminés.
Le choix du niveau d’expression va varier selon les espèces. Il dépend :
- de la plus ou moins grande variabilité des pertes d’azote consécutive à l’utilisation digestive ou à l’utilisation métabolique ;
- de la possibilité de connaître ou d’estimer assez précisément ces pertes selon les aliments ou les caractéristiques des animaux.
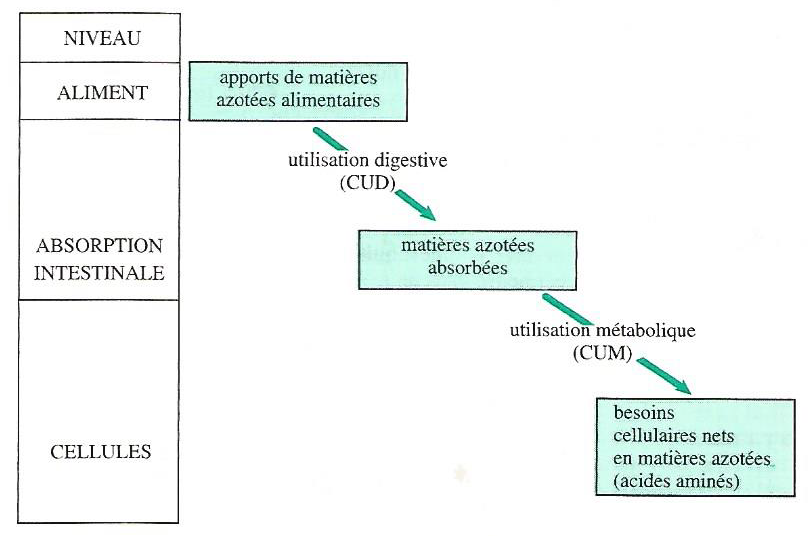
Figure 4.10 Niveaux d’expression des apports et des besoins azotés
4.4.6.1 Chez les monogastriques
Chez les monogastriques stricts, l’utilisation digestive varie peu et l’utilisation métabolique dépend principalement de l’apport en acides aminés essentiels. Le niveau retenu est le niveau de l’apport ou celui de l’absorption. La valeur des aliments et les besoins sont exprimés en Matières Azotées Totales ou Protéines Brutes, en Acides Aminés Essentiels ou AA Essentiels Digestibles.
Les monogastriques sont tributaires des acides aminés alimentaires et plus particulièrement des acides aminés indispensables. Ceux-ci doivent donc se trouver en quantité suffisante au niveau cellulaire simultanément avec l’énergie nécessaire à la réalisation des synthèses protéiques.
LE CAS DU PORC
Digestibilité iléale chez le porc
Donc, chez le porc, les apports recommandés sont exprimés en MAT et en AA Intestinaux digestibles, rapportés à l’énergie nette (EN) : \(AAID/MJ~d’EN~~ou~~AAID/kcal~d’EN\). On mesure la digestibilité à la sortie de l’intestin grêle, c’est-à-dire au niveau de l’iléon. En effet, la microflore présente dans le caecum et le gros intestin peut dégrader les matières azotées résiduelles, non dégradées dans l’intestin grêle, en ammoniac. La microflore peut aussi remanier ces matières azotées résiduelles pour générer d’autres acides aminés. Or, les protéines ou Acides aminés ne sont plus absorbés après l’intestin grêle et donc, ce qu’on va retrouver dans les fèces ne correspondra pas à l’utilisation digestive effective. Alors, on évalue la digestibilité iléale.
Utilisation de l’azote chez le porc et impact sur l’environnement
On considère que le porc ne fixe qu’un tiers (\(1/3\)) de l’azote ingéré. Les deux autres tiers (\(2/3\)) sont rejetés dans l’environnement. On estime que 50 % de ces rejets sont volatilisés sous forme d’ammoniac (\(NH3\)) et les 50 % restants retourneraient au sol après épandage des lisiers au champ et serait utilisés par les plantes à condition que les quantités (doses) d’épandage soient respectées.
Les émissions d’azote constituent une perte pour l’agriculteur et ont des conséquences sur la santé humaine et le milieu naturel. Il est donc important de maîtriser au mieux l’alimentation porcine pour réduire au maximum les rejets d’azote.
Amélioration de l’équilibre azoté des rations chez le porc
La qualité ou l’équilibre azoté d’une ration passe par :
- le respect des apports alimentaires et l’équilibre en acides aminés de la ration ;
- l’adaptation des aliments au type de production ou au stade physiologique.
Pour couvrir les besoins azotés on associe différentes matières premières (qui doivent apporter à l’animal les acides aminés essentiels) et une quantité suffisante de protéines (garantissant l’apport d’une quantité globale minimale de l’ensemble des acides aminés non indispensables).
Les céréales qui sont les éléments de base des rations des monogastriques, présentent plusieurs facteurs limitant en acides aminés indispensables alors que les tourteaux, les oléagineux et les oléo-protéagineux en sont relativement bien pourvus. Le tourteau de soja est le seul à présenter un taux élevé en lysine. Ainsi, l’association céréales et tourteaux permet de couvrir les besoins en acides aminés indispensables, mais avec des taux azotés relativement élevés, voire excessif, ce qui entraîne un gaspillage inutile avec l’augmentation d’azote dans les effluents d’élevage et des risques accrus de pollution.
Pour pallier ce problème, on peut supplémenter avec des acides aminés de synthèse.
Le concept de protéine idéale chez le porc
La protéine idéale est la protéine dont l’équilibre en acides aminés essentiels permet la protéosynthèse maximale. Ces acides aminés essentiels doivent présenter des rapports constants entre eux en fonction de la composition en acides aminés des protéines déposées (synthétisées).
La lysine est le plus souvent le facteur limitant primaire. Ainsi, on caractérise la protéine idéale en exprimant les apports recommandés en acides aminés essentiels par référence à la lysine à laquelle on attribue arbitrairement la valeur 100 (Tableau 4.5).
Tableau 4.5 Rapport recommandé entre les besoins en acides aminés indispensables chez le porc en croissance par référence à la lysine
| Acides aminés | Rapports recommandés |
|---|---|
| Lysine | 100 |
| Thréonine | 60 – 65 |
| Tryptophane | 19 |
| Méthionine + cystine | 60 |
| Méthionine | 30 |
| Isoleucine | 60 |
| Leucine | 72 |
| Valine | 70 |
| Histidine | 26 |
| Phénylalanine + tyrosine | 100 |
| Arginine | 45 |
Il est souhaitable que l’aliment reflète les proportions en tenant compte des rendements d’utilisation des acides aminés. La protéine idéale n’a pas la même composition selon qu’elle concerne le porc en croissance, la truie gestante ou la truie allaitante. La formulation des aliments doit aussi avoir pour objectif de se rapprocher de la protéine idéale afin de permettre une utilisation maximale des matières azotées ingérées et par conséquent, une réduction des rejets dans les lisiers.
LE CAS DES VOLAILLES
Il est difficile d’estimer les besoins des volailles car il existe de nombreuses interactions entre plusieurs facteurs : le génotype, l’environnement (température, lumière, vitesse de l’air, …) et les conditions d’élevage. Cependant, on retrouve également un concept de protéine idéale, en particulier chez le poulet, basé sur les acides aminés essentiels digestibles (Tableau 4.6).
Tableau 4.6 Rapport recommandé entre les besoins en acides aminés indispensables chez le poulet par référence à la lysine
| Acides aminés | Rapports recommandés |
|---|---|
| Lysine | 100 |
| Thréonine | 73 |
| Tryptophane | 18 |
| Acides aminés soufrés | 70 |
| Valine | 82 |
| Isoleucine | 73 |
| Arginine | 114 |
LA NOTION D’INDICE DE CONSOMMATION (IC)
C’est un indicateur qui permet de mesurer l’impact de l’alimentation sur les performances zootechniques. Autrement dit : est-ce que l’animal valorise bien sa ration ?
Par définition, c’est le rapport entre la quantité d’aliment consommée et une quantité de croît. Par exemple, dire qu’un animal, à une phase de croissance donnée, a un IC = 1,2 signifie qu’il faut 1,2 kg de son aliment pour croître de 1 kg en poids vif. L’IC est aussi appelé taux de conversion de l’aliment.
EN RÉSUMÉ
Chez les porcs et les volailles, la valeur azotée des aliments et les apports alimentaires recommandés sont exprimés au niveau de l’aliment, c’est-à-dire en matières azotées totales (MAT) ou protéines brutes (PB).
Pour couvrir le besoin azoté du monogastrique, il est nécessaire d’apporter les acides aminés essentiels et une certaine quantité de protéines couvrant le besoin en acides aminés banals.
La supplémentation consiste à couvrir les besoins en acides aminés indispensables avec le minimum de protéines naturelles et un ou plusieurs acides aminés de synthèse. L’amélioration de l’équilibre en acides aminés ainsi obtenu permet une réduction du taux azoté de l’aliment, donc une économie.
4.4.6.2 Chez les ruminants
Chez les ruminants, on ne peut pas considérer le niveau de l’apport car les matières azotées contenues dans l’aliment sont en partie remaniées (dégradées) par les bactéries du rumen-réseau (cf. chapitre digestion). La valeur des aliments et les besoins sont exprimés en protéines digestibles dans l’intestin (\(PDI\)), ou en acides aminés digestibles intestinaux (\(AADI\)) pour les vaches laitières.
Chez les ruminants, les acides aminés absorbés au niveau de l’intestin ont une double origine : alimentaire et microbienne :
- les acides aminés alimentaires sont issus des protéines ingérées et non dégradées dans le rumen ;
- les acides aminés microbiens sont issus des protéines microbiennes formées dans le rumen.
De ce fait, le ruminant peut :
- synthétiser des acides aminés indispensables. Il n’est pas obligatoire de les lui fournir dans son alimentation, surtout si les besoins sont peu élevés ;
- utiliser d’autres sources azotées que les protéines (l’urée par exemple).
UTILISATION DIGESTIVE DES MATIÈRES AZOTÉES
Dans le rumen-réseau
La dégradation des matières azotées dans le rumen est une protéolyse microbienne dont le produit est essentiellement l’ammoniac. Cette dégradabilité dépend :
- de la structure chimique des matières azotées (les protéines sont moins dégradables) ;
- de leur accessibilité par les micro-organismes ;
- d’éventuels traitements technologiques ;
- de l’activité microbienne dans le rumen.
Les bactéries utilisent l’ammoniac (issu de la dégradation des matières azotées dans le rumen) pour fabriquer leurs propres protéines. Pour réaliser cette synthèse, ces bactéries ont aussi besoin d’énergie et de chaînes carbonées. Ces deux éléments sont apportés par les glucides fermentescibles.
En conditions idéales, l’ammoniac est utilisé pour la synthèse de matières azotées microbiennes. En conditions défavorables (excès d’ammoniaque ou déficit en glucides fermentescibles), il y a trop d’ammoniac pour la capacité de protéosynthèse microbienne. L’ammoniac s’accumule dans le rumen et est absorbé. Capté par le foie, il est transformé en urée qui est excrétée par le rein. C’est donc une perte. De plus, lorsque le taux d’ammoniaque du sang devient excessif, des troubles hépatiques et des accidents de toxicité peuvent apparaître.
La dégradation des matières azotées est estimée à l’aide de tests de dégradabilité qui permettent d’obtenir la dégradabilité théorique ou DT. Les matières azotées non dégradables sont quantifiées par : \(1,11 * MAT * (1 – DT)\).
Après le rumen-réseau
Les matières azotées qui quittent le rumen-réseau sont :
- les matières azotées alimentaires non dégradées, constituées uniquement de protéines ;
- les matières azotées microbiennes issues de la protéosynthèse ;
- un peu d’ammoniac non utilisé et non absorbé (2 à 5 % de l’azote total) ;
- une petite quantité de matières azotées endogènes (sécrétions enzymatiques).
L’azote microbien représente en général nettement plus de la moitié du flux total d’azote entrant dans le duodénum (50 à 90 %).
On estime que, chez une vache laitière produisant \(25~kg\) de lait et recevant une ration mixte classique (foin + concentré), les microorganismes fabriquent \(2~kg\) de matière sèche microbienne par jour, dont 50 % sous forme de protéines, soit \(1~kg\) de protéines microbiennes.
Sous l’action des enzymes protéolytiques et des sucs digestifs, toutes les protéines, d’origine alimentaire ou microbienne, fournissent des acides aminés qui seront absorbés.
Transformations subies par les matières azotées après le rumen-réseau
- Dans la caillette et l’intestin grêle
Sous l’action des enzymes protéolytiques des sucs digestifs, toutes les protéines, d’origine alimentaire ou microbienne, fournissent des acides aminés.
- Dans le gros intestin
On peut négliger ce qui se passe dans le gros intestin. Il y a une reprise de l’activité microbienne avec protéolyse et synthèse microbienne, mais la quantité d’acides aminés absorbés à ce niveau est très faible.
UTILISATION MÉTABOLIQUE DES ACIDES AMINÉS ABSORBÉS
Les besoins en acides aminés des celJules des tissus des ruminants sont les mêmes que dans le cas des monogastriques : les acides aminés arrivant au niveau des cellules doivent, pour être utilisés au mieux, figurer dans des proportions très voisines de celles des besoins des tissus. Les principes de base de l’alimentation azotée des monogastriques restent valables :
il existe des acides aminés indispensables et des acides aminés non indispensables ;
l’efficacité de l’apport des acides aminés pour couvrir les besoins cellulaires peut également être estimée par le coefficient d’utilisation métabolique réel.
Les acides aminés ont une double origine chez le ruminant :
une origine alimentaire, la digestion des protéines non dégradées dans le rumen (\(AAa\)), et
une origine microbienne, la digestion des protéines microbiennes provenant de la protéosynthèse (\(AAm\)).
La relation entre la composition en acides aminés des matières azotées ingérées et celle du mélange d’acides aminés absorbés dans l’intestin est faible.
LE SYSTÈME D’EXPRESSION DE LA VALEUR AZOTÉE DES ALIMENTS CHEZ LE RUMINANT (PDI)
L’abréviation PDI est la contraction d’une expression plus exacte et plus explicite : Protéines vraies réellement Digestibles dans l’Intestin grêle. Le système PDI exprime les apports alimentaires et les besoins des animaux au niveau de l’absorption des acides aminés dans l’intestin grêle.
La valeur azotée d’un aliment exprimée en PDI est la somme de deux fractions :
les protéines réellement digestibles dans l’intestin grêle d’origine alimentaire ou \(PDIA\). Ce sont les acides aminés réellement absorbés provenant des protéines alimentaires non dégradées dans le rumen ;
les protéines réellement digestibles dans l’intestin grêle d’origine microbienne ou \(PDIM\). Ce sont les acides aminés réellement absorbés provenant des protéines microbiennes issues de la protéosynthèse.
La valeur \(PDI\) de l’aliment est donc : \(PDI = PDIA + PDIM\), c’est-à-dire que la somme des acides aminés absorbés au niveau de l’intestin (\(AAa + AAm\)).
En tenant compte des deux principaux facteurs limitants de la protéosynthèse que sont l’énergie fermentescible et les matières azotées dégradables, on peut distinguer deux valeurs \(PDIM\) :
la quantité de protéines microbiennes qui peut être obtenue à partir de l’énergie fermentescible disponible est la valeur \(PDIME\) de l’aliment (Protéines Digestibles dans l’Intestin d’origine Microbienne permise par l’Energie) ;
la quantité de protéines microbiennes, qui peut être obtenue à partir de l’azote dégradable disponible est la valeur \(PDIMN\) de l’aliment (protéines digestibles dans l’intestin d’origine microbienne permise par l’azote).
Chaque aliment a donc deux valeurs azotées PDI :
\(PDIA + PDIMN = PDIN\) (Protéines Digestibles dans l’Intestin permises par l’azote),
\(PDIA + PDIME = PDIE\) (Protéines Digestibles dans l’Intestin permises par l’Energie).
La plus petite de ces deux valeurs est la valeur azotée effective de l’aliment distribué seul. La plus élevée peut être considérée comme une valeur potentielle, maximale, qui n’est atteinte que si l’aliment est associé avec d’autres aliments dont le facteur limitant est différent.
La valeur \(PDIN\) représente la valeur \(PDI\) d’un aliment s’il est inclu dans une ration déficitaire en azote dégradable. La valeur \(PDIE\) représente la valeur \(PDI\) d’un aliment s’il est inclus dans une ration où l’énergie est le facteur limitant de la synthèse microbienne.
Pour une ration, on ne peut pas additionner des valeurs \(PDIN\) et des valeurs \(PDIE\). La valeur pour une ration est obtenue en faisant séparément la somme des \(PDIN\) et la somme des \(PDIE\) des aliments, et en retenant la plus faible, de sorte que la valeur réelle d’un aliment peut être \(PDIN\) ou \(PDIE\) selon le contexte (Figure 4.11).
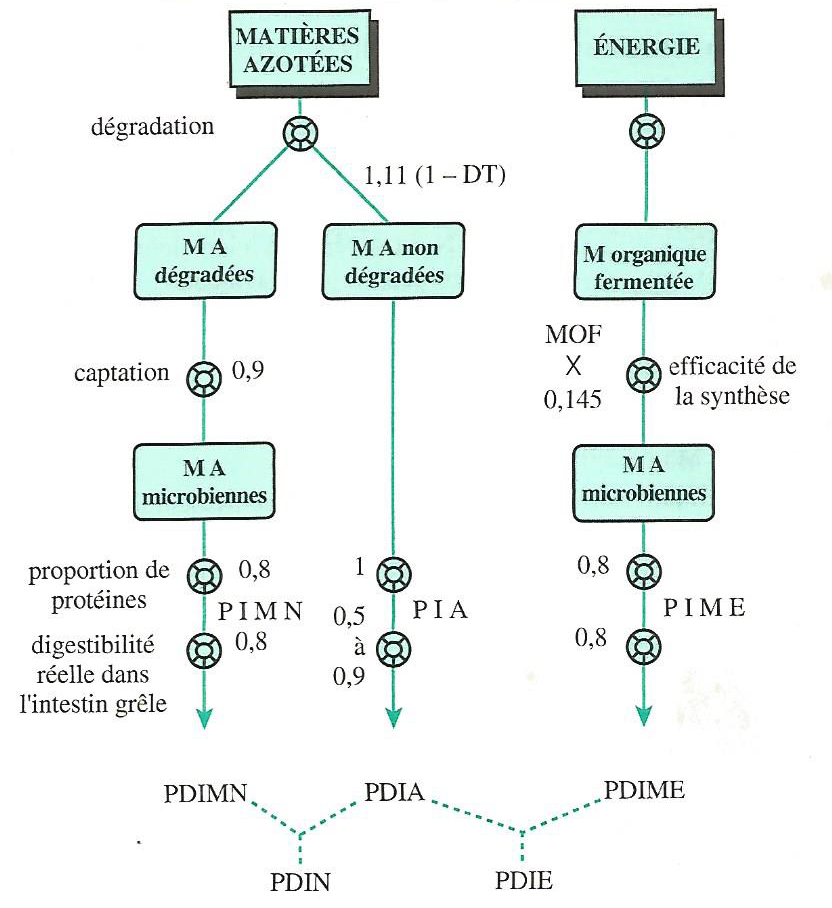
Figure 4.11. Principe de la détermination des valeurs PDI des aliments
L’association judicieuse des aliments dans la ration permet de bénéficier de leur complémentarité. L’énergie fermentescible des uns permet de valoriser les matières azotées dégradables des autres. L’optimum est obtenu lorsque les différents aliments peuvent atteindre leur valeur azotée maximale.
L’intérêt du système \(PDI\) est donc d’obtenir, pour un mélange d’aliments dans une ration, l’égalité :
\[Somme~des~apports~PDIN = Somme~des~apports~PDIE\]
La plupart des aliments ont des valeurs \(PDIN\) et \(PDIE\) différentes. On peut distinguer :
les aliments dont la valeur \(PDIE\) est supérieure à la valeur \(PDIN\) : pailles, fourrages de mauvaise qualité, racines, pulpes, ensilages de maïs, céréales, etc. Les matières azotées dégradables sont alors le facteur limitant de la protéosynthèse. Ce sont en outre tous des aliments dont le taux azoté est faible (MAT < 12% de la MS),
les aliments dont la valeur \(PDIN\) est supérieure à la valeur \(PDIE\) : certains fourrages récoltés jeunes, ou à base de légumineuses, les protéagineux, les tourteaux et quelques sous-produits comme les drêches. L’énergie fermentescible est alors le facteur limitant de la protéosynthèse. Ils ont en outre tous un taux élevé de matières azotées (MAT > 14 % de la MS).
L’urée est un cas particulier. Elle n’apporte pas d’énergie et n’a donc qu’une valeur \(PDIN\) : 1472 g/kg de MS (ou 1443 g/kg ? je vais vérifier …).
Un des objectifs du rationnement est de réaliser une association judicieuse d’aliments à partir de chacune de ces deux catégories. Les deux exemples des Tableau 4.7 et 4.8 montrent qu’il est possible, à partir de deux aliments aux caractéristiques différentes, d’obtenir une combinaison permettant l’égalité voulue (\(Somme~des~apports~PDIN = Somme~des~apports~PDIE\))
Tableau 4.7 Valeurs PDI et valeur azotée de deux aliments (ensilage de maïs et ensilage d’herbe) et de trois associations de ces deux aliments
| PDIN (en g) | PDIE (en g) | Valeur azotée (g de PDI) | ||
|---|---|---|---|---|
| 1 | Ensilage de maïs | 52 | 66 | 52 |
| 2 | 80 % ensilage de maïs + 20 % ensilage d’herbe | 57 | 65 | 57 |
| 3 | 50 % ensilage de maïs + 50 % ensilage d’herbe | 65 | 65 | 65 |
| 4 | 30 % ensilage de maïs + 70 % ensilage d’herbe | 70 | 64 | 64 |
| 5 | Ensilage d’herbe | 78 | 63 | 63 |
Tableau 4.8 Valeurs PDI et valeur azotée de deux aliments (orge et tourteau de soja) et de trois associations de ces deux aliments
| PDIN (en g) | PDIE (en g) | Valeur azotée (g de PDI) | ||
|---|---|---|---|---|
| 1 | Orge | 79 | 102 | 79 |
| 2 | 90 % orge + 10 % tourteau de soja | 108 | 117 | 108 |
| 3 | 84 % orge + 16 % tourteau de soja | 126 | 126 | 126 |
| 4 | 50 % orge + 50 % tourteau de soja | 225 | 178 | 178 |
| 5 | Soya | 371 | 254 | 254 |
4.4.7 La complémentation azotée des rations dans le système \(PDI\)
Dans une ration, l’optimum est d’obtenir un équilibre entre les apports \(UF\), \(PDIN\) et \(PDIE\). Une alimentation azotée correcte doit satisfaire les besoins azotés des microorganismes, tout en couvrant les besoins du ruminant. C’est ce que le système \(PDI\) traduit, au niveau d’une ration équilibrée, par l’égalité \(PDIN = PDIE\). Mais il est difficile d’obtenir une égalité stricte. Ne pas trop s’en écarter est un objectif plus réaliste.
Dans la pratique, on vérifie l’équation du seuil PDI suivante : \(seuil~PDI = \frac{PDIN-PDIE}{UF}\). Ces valeurs seuils sont disponibles dans les tables INRA 2007.
CAS PARTICULIER DES LAITIÈRES FORTES PRODUCTRICES
Chez les vaches laitières à haut niveau de production, les synthèses microbiennes ne permettent pas de couvrir les besoins de l’animal en acides aminés indispensables. Il est donc nécessaire de les apporter dans l’alimentation. Il faudra protéger ces matières azotées pour éviter que les microorganismes ne les dégradent. Cette protection est assurée par des traitements technologiques (exemple avec le tannage des tourteaux.
EN RÉSUMÉ CHEZ LE RUMINANT
Les ruminants adultes ont les mêmes besoins cellulaires en acides aminés que les monogastriques, c’est-à-dire des besoins en acides aminés indispensables et en acides aminés banals. Mais les acides aminés absorbés au niveau de l’intestin grêle ont 2 origines : alimentaire et microbienne.
Chez les ruminants adultes, les apports et les besoins sont exprimés en protéines digestibles dans l’intestin (PDI). La valeur PDI des aliments est la somme de 2 fractions (\(PDI = PDIA + PDIM\)) :
- \(PDIA\), protéines digestibles intestinales d’origine alimentaire ;
- \(PDIM\), protéines digestibles intestinales d’origine microbienne.
La valeur \(PDIM\) dépend de la disponibilité dans le rumen des 2 facteurs limitants principaux de la protéosynthèse : l’énergie fermentescible et les matières azotées dégradables. La teneur en \(PDIM\) d’un aliment est alors caractérisée par 2 valeurs dépendant :
- l’une, de la teneur en énergie fermentescible, \(PDIME\), protéines digestibles intestinales microbiennes permises par l’énergie ;
- l’autre, de la teneur en matières azotées dégradables, \(PDIMN\), protéines digestibles intestinales microbiennes permises par l’azote.
Les aliments ont donc 2 valeurs PDI : 1. PDIN = PDIA + PDIMN 1. PDIE = PDIA + PDIME
4.5 Expression des besoins et des apports en minéraux et en vitamines
Comme c’est le cas pour l’énergie et les matières azotées, les apports en minéraux et en vitamines des différents aliments de la ration doivent être évalués et additionnés, puis comparés aux besoins de l’animal. Si les besoins ne sont pas couverts, un apport complémentaire en minéraux et en vitamines est alors réalisé, généralement sous la forme d’un complexe minéral-vitaminé (CMV), dont la composition sera choisie en fonction des déficits existants.
Cette démarche suppose de connaître préalablement d’une part, la teneur en minéraux et en vitamines des aliments de la ration, et d’autre part les besoins de l’animal. Les apports en minéraux des aliments sont exprimés :
- en \(g/kg\) de MS d’aliment pour les macro-éléments (Ca, P, K, Na, Cl, S et Mg) ;
- en \(mg/kg\) de MS d’aliment ou en \(ppm\) pour les oligo-éléments (Fe, Se, Zn, Cu, I, Co, Mn).
Les apports en vitamines sont quant à eux exprimés en \(mg/kg\) de MS d’aliment ou en \(UI/kg\) de MS d’aliment.
ppm (partie par million) : unité de mesure désignant une concentration d’une substance égale à un millionième : par exemple des mg/kg, des g/tonne, …
UI (Unité Internationale) : unité de mesure permettant d’évaluer la quantité d’une substance sur base de son activité biologique. L’UI diffère donc d’une substance à l’autre. Ainsi, 1 UI de vitamine \(A = 0,33~µg\) de vitamine A et 1 UI de vitamine $D = 0,025~µg de vitamine D.*
Les modalités d’expression des besoins en minéraux et en vitamines sont différentes selon que l’on s’intéresse aux macro-éléments et aux vitamines d’une part, ou aux oligo-éléments d’autre part.
Pour les macro-éléments et les vitamines :
les besoins de l’animal sont exprimés en termes de besoins absolus, c’est-à-dire en \(g/jour\) (ou parfois en \(UI/jour\) pour les vitamines). Par exemple, les besoins en Ca d’une vache laitière de \(650~kg\) produisant \(25~L\) sont de \(107,5~g/jour\). Ses besoins en vitamine A sont quant à eux de \(90000~UI/jour\).
Pour les oligo-éléments il en va différemment :
Les besoins en oligo-éléments des animaux ne sont en réalité pas connus avec précision. Aussi, ils sont exprimés en termes de besoins relatifs, et font l’objet de recommandations à suivre quant à la teneur en oligo-éléments à atteindre dans la MS de la ration de l’animal, avec fixation d’un seuil de carence et d’un seuil de toxicité.
Les besoins en oligo-éléments sont donc exprimés en \(ppm\) ou en \(mg/kg\) de MS ingérée. Par exemple, les besoins en Cu d’une vache laitière de \(650~kg\) produisant \(25~L\) sont de \(8~à~10~mg/kg\) de MS de ration ou \(8~à~10~ppm\).
En termes d’apports alimentaires, il faut retenir que pour les minéraux, les teneurs des fourrages sont assez variables, alors que celles des aliments concentrés sont relativement stables. Par conséquent, il est toujours souhaitable de réaliser une analyse de la composition chimique de son fourrage, alors que pour les aliments concentrés, l’utilisation des tables de composition permet une estimation assez fiable des teneurs.
4.3.5.1 Comment choisir un système d’expression de l’énergie
On rappelle que la valeur énergétique des aliments distribués et les besoins (énergétiques) des animaux doivent être exprimés dans le même système d’unité. Le problème est de choisir le niveau de l’utilisation de l’énergie auquel on se situe.
C’est l’étape ultime de la transformation d’un aliment par l’animal qui constitue le niveau le plus pertinent pour estimer la valeur énergétique de cet aliment. L’énergie nette correspondant au niveau cellulaire est préférable, car elle tient compte de toutes les catégories de pertes énergétiques intervenant entre l’ingestion et l’utilisation par la cellule.
La principale difficulté d’un système fondé sur l’énergie nette est l’estimation du rendement k d’utilisation de l’énergie métabolisable en énergie nette :
en raison des variations du rendement k en fonction des caractéristiques de la ration et de la fonction physiologique,
parce qu’au moins deux fonctions physiologiques coexistent chez un animal (entretien et au moins une production) et que le rendement global de l’énergie métabolisable en énergie nette dépend de l’importance relative de l’entretien et de la production (revoir Figure 4.2).
Il y a donc un grand nombre de situations possibles et en pratique, le choix d’un système basé sur l’énergie nette rend nécessaire de choisir une situation « standard » représentative d’un maximum de cas.
En pratique, on peut distinguer deux grands groupes d’animaux :
pour les animaux qui reçoivent des aliments de nature et de composition très différentes, conduisant de ce fait à des pertes énergétiques très variables, on utilise un système basé sur l’énergie nette (EN). C’est le cas des ruminants et des chevaux. Pour les ruminants, on dispose en outre d’informations précises relatives au rendement k.
pour les animaux qui ingèrent des rations moins hétérogènes (nature et composition pas très différentes) et qui ont des pertes organiques (EF, EG, EU) et d’extra-chaleur moins variables, on utilise des systèmes fondés sur l’énergie métabolisable (EM) ou l’énergie digestible (ED). C’est le cas :
Cependant, pour les porcs, les systèmes utilisés dans différents pays, énergie digestible (ED), énergie métabolisable (EM) et énergie nette d’engraissement (ENE), font varier le classement des matières premières.